Обмен веществ (метаболизм), совокупность химических процессов, обеспечивающих жизнедеятельность организма. Химические превращения в
организме осуществляются в двух противоположных направлениях - синтез сложных соединений из более простых (анаболизм, или ассимиляция) и расщепление сложных
соединений до более простых (катаболизм, или диссимиляция).
Первоначальные представления об обмене веществ возникли в связи с изучением процессов обмена между организмами и внешней средой (так называемый
внешний, или общий, обмен веществ). Последние исследования превращений веществ внутри организма привели к представлениям о внутреннем, или
промежуточном, обмене веществ.
Во внутреннем и внешнем обмене веществ принято различать структурный (пластический) и энергетический обмены. В структурном обмене рассматривают
превращения различных соединений в организме, их перенос (транспорт) внутри организма и между организмом и средой. В энергетическом обмене рассматривают
превращения химической энергии, образующейся в обмена веществ, в тепло, мышечную работу, а также механизмы ее использования в активном транспорте,
биосинтезе и др.
В соответствии с природой участвующих в обмене веществ соединений различают органический обмен (обмен углеводов, липидов, азотсодержащих соединений
и др.) и минер. обмен (водно-солевой обмен и обмен микроэлементов); в физиологии выделяют также газовый обмен. Обмен веществ с участием свободного
О2 наз. аэробным, без участия О2 - анаэробным. Ввиду различий обмена веществ у организмов, принадлежащих к различным
таксономическим группам, выделяют обмен веществ растений, животных, микроорганизмов, а также более мелких таксономических единиц, например обмен
веществ млекопитающих, злаковых, дрожжей, человека, бактерии Escherichia coli. При изучении обмена веществ учитывают половые и возрастные
различия, а также отклонения в обмене веществ, вызванные влиянием внешней среды и питания. Раздельно рассматривают обмен веществ в различных
тканях и органах. Устойчивые отклонения обмена веществ от нормы квалифицируют как болезни обмена веществ
Структурный обмен
В зависимости от того, в какой химической форме живые организмы способны усваивать из внешней среды углерод, они делятся на две большие группы - автотрофы
и гетеротрофы. Для первых основным источником углерода служит СО2, для вторых-разлагаемые органические соединения. Автотрофное питание
осуществляют зеленые растения и фотосинтезирующие бактерии, гетеротрофное - животные и грибы. У микроорганизмов встречаются тот и другой типы питания.
обмен веществ автотрофных организмов является по преимуществу анаболическим, гетеротрофных - катаболическим. Основу пластического обмена составляет
органический обмен. Традиционное разделение его на углеводный обмен, липидный обмен и обмен азотсодержащих соединений обусловлено большой распространенностью
в живой природе соединений этих классов и различием их свойств.
Субстратами органического обмена являются вещества, поступающие из внешней среды, и вещества внутреннего происхождения. В процессе обмена веществ
часть конечных продуктов выводится во внешнюю среду, другая часть используется организмом. Конечные продукты органического обмена в тканях, способные
накапливаться или расходоваться в зависимости от условий существования организма (например, триацилглицерины, гликоген, крахмал, проламины), называются
запасными, или резервными, веществами. Если скорость поглощения субстратов превосходит скорость выведения конечных продуктов, то анаболизм преобладает над
катаболизмом и организм развивается или накапливает резервные вещества. При равенстве этих скоростей рост организма прекращается и обмен веществ
преходит в состояние, близкое к стационарному. В случае превышения скорости выведения конечных продуктов над скоростью потребления после истощения запаса
рзервных веществ организм обычно погибает. Последнее наблюдается при искусственном ограничении потребления внешних субстратов (например, алиментарная
дистрофия при голодании животных, самосбраживание дрожжей в условиях дефицита углеводов) или в естественных условиях (например, при интенсивном дыхании
плодов и семян растений).
Главный катаболический процесс в обмен евеществ-биологическое окисление (совокупность реакций окисления, протекающих во всех живых клетках, -
дыхание и окислительное фосфорилирование). Интегральной характеристикой
биологического окисления служит дыхательный коэффициент, который представляет собой отношение объема выделенного организмом СО2 к объему
поглощенного О2. При окислении углеводов объем расходуемого О2 соответствует объему образующегося СО2 и поэтому дыхательный
коэффициент в этих случаях равен единице. При окислении жиров и белков такое соответствие отсутствует, т.к. кроме окисления углерода до СО2 часть
О2 расходуется на окисление водорода с образованием воды. Вследствие этого величины дыхательного коэффициента в случае окисления жиров и белков
составляют соответственно около 0,7 и 0,8. Подавляющая часть белкового азота при окислении белка в организме млекопитающих переходит в мочевину. Поэтому по
дыхательному коэффициенту и данным о количестве выделяемой мочевины у млекопитающих можно определять соотношение участвующих в биологическом окислении
углеводов, жиров и белков.
Основным анаболическим процессом, противоположным биологическому окислению, является осуществляемый автотрофами фотосинтезорганических соединений из СО2 и воды. Вместе с тем автотрофы
осуществляют частичное окисление продуктов фотосинтеза. Для характеристики их общего обмена веществ также используют дыхательный коэффициент.
Последовательности реакций в организме, в которых осуществляется превращение субстратов в конечные продукты обмена веществ, наз. путями обмена
веществ, или метаболическими путями, а вещества, участвующие в этих реакциях, - метаболитами. В зависимости от характера превращения субстратов
метаболические пути подразделяют на анаболические и катаболические. Обратимые участки метаболических путей, состоящие из равновесных реакций и используемые
организмами как для синтеза, так и для расщепления сложных соединений, называются амфиболическими. Подавляющую часть реакций, составляющих метаболические
пути, катализируют ферменты. Для своего функционирования многие ферменты нуждаются в низкомолекулярных соединениях, называемых коферментами. У высших животных большая часть коферментов (или их непосредственных
предшественников) поступает в организм с пищей в виде незаменимых факторов питания-витаминов.
В простых случаях стационарность обмена веществ обеспечивают метаболические пути, образованные линейными последовательностями реакций (например,
гликолиз, синтез и расщепление жирных кислот). Если метаболические пути включают реакции, в которых образуются вещества, не выводимые во внешнюю среду, то
стационарность обмена веществ поддерживается дополнительными реакциями, обеспечивающими регенерирование этих веществ в предшествующие метаболиты. В
результате метаболические пути принимают вид циклических последовательностей реакций (см., например, трикарбоновых кислот цикл, глиоксилатный цикл).
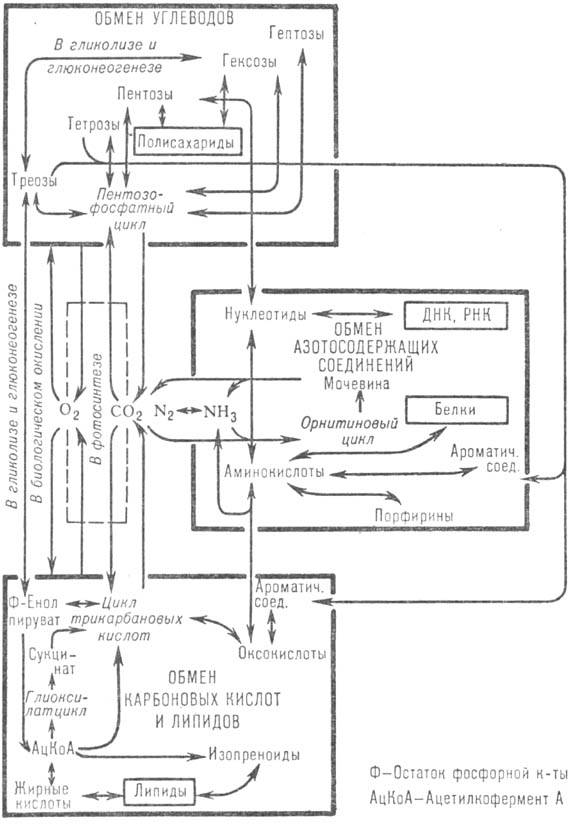
Различные метаболические пути включают много общих промежуточных соединений и поэтому образуют единую сеть реакций, которая обнаруживает структурную
регулярность. Эта регулярность, обусловленная наличием большого количества сходных реакций у метаболитов с одинаковыми функциональными группами, отражена в
семах 1-3. Эти схемы объединяют данные по обмену веществ у животных, растений и микроорганизмов. Реакции обмена веществ человека и близких по
обмену веществ млекопитающих выделены жирными стрелками. Соединения с одинаковыми функциональными группами размещены на схемах на одной горизонтали в
порядке увеличения числа атомов С в скелете их молекул. По вертикали схемы разделены на участки, связанные сходными реакциями и включающие соединения с
одинаковым числом атомов С в скелете. Эти участки, соответствующим образом пронумерованные, названы периодами (номер периода соответствует числу атомов С в
скелете молекулы). Смежные периоды имеют сходную структуру; соединения, занимающие в них соответствующее положение, являются гомологами. Названия и формулы
некоторых кислот, встречающихся в тексте и на схемах 1-3, приведены в таблице.
Обмен углеводов. Важнейший метаболический путь в обмене углеводов - гликолиз,
в котором осуществляется превращение гексоз в две молекулы лактата. Этот путь широко представлен в тканях животных и обеспечивает двигательную функцию
скелетных мышц. В печени амфиболические реакции этого пути участвуют в глюконогенезе - биосинтезе глюкозы из образующегося в мышцах лактата. Гликолиз
рассматривают в качестве основного пути, связывающего углеводный обмен с обменом карбоновых кислот и липидов. К модификации гликолиза относят путь спиртового
брожения. В отсутствие свободного О2 дрожжи по этому пути
количественно расщепляют глюкозу на этанол и СО2, осуществляя так называемое анаэробное дыхание.
Названия и формулы некоторых кислот, встречающихся в тексте и на схемах 1-3
Название ионизир. Название кислоты Формула формы кислоты
Важную роль в катаболизме углеводов играет пентозофосфатный цикл. Ключевые
реакции этого пути - окисление глюкозо-6-фосфата до 6-фосфоглюконата и декарбоксилирование последнего с образованием СО2, воды и
рибулозо-5-фосфата. Благодаря цикличности этого процесса обеспечивается стационарность окисления глюкозы в тканях. Так же как и в случае гликолиза,
равновесные реакции этого пути составляют амфиболический участок, который наряду с реакцией карбоксилирования рибулозо-1,5-дифосфата обеспечивает при
фотосинтезе у зеленых растений обратный процесс - биосинтез глюкозы из СО2 и воды. При этом глюкоза в результате ферментативного превращ. в олиго-
и полисахариды выводится из сферы обмена веществ в виде крахмала, целлюлозы и др.
С реакциями пентозофосфатного цикла связан метаболизм входящих в состав нуклеиновых кислот пентоз, а также биосинтез углеводных предшественников
биополимера лигнина и ароматических аминокислот.
Основным углеводным субстратом в обмене веществ человека и высших животных служит глюкоза. Она сохраняется в виде резервного полисахарида гликогена
в печени и частично в мышцах. Восстановление запасов гликогена происходит благодаря его синтезу из глюкозы, образуемой при глюконеогенезе или поступающей в
кровоток через стенки кишечника. В последний глюкоза попадает в результате гидролиза крахмала пищевых продуктов амилазой слюны и ферментами
желудочно-кишечного тракта.
Наряду с крахмалом высшие животные усваивают гликоген, некоторые олигосахариды и дисахариды, например сахарозу, мальтозу, лактозу и др.
Специализирированные микроорганизмы могут расщеплять ксилан, целлюлозу, хитин, лигнин и др. устойчивые полисахариды. Способность жвачных животных усваивать
целлюлозу и ксилан обусловлена жизнедеятельностью микрофлоры, обитающей в сложном желудке животных.
На схеме 1 показаны основные пути обмена углеводов. В ее верхей части представлены моносахариды и их производные с трео-конфигурацией в положении
3-4. В нижней части размещены ряды соединения тех же классов, но с эритро-конфигурацией в этом же положении. Высшие моносахариды образуются из низших
в обратимых реакциях конденсации с соединениями, содержащими в молекуле два или три атома С. В реакциях, катализируемых транскетолазами и соединяющих периоды
n и n + 2 схемы (начиная с n = 2), из D-альдоз нижнего ряда образуются 3,4-D-трео-кетозы верхнего ряда, имеющие углеродный скелет
на два атома С больше, чем исходные альдозы. В реакциях конденсации D-альдоз с соединениями, содержащими три атома С, обеспечивающих связь между периодами
n и n + 3, углеродный скелет увеличивается на 3 атома С. Пентозы образуются в результате декарбоксилирования уридиндифосфатных (УДФ) поизводных
уроновых кислот. Переход от 3,4-D-тpeo-моносахаридов к 3,4-D-эритро-моносахаридам (периоды 5 и 6) осуществляется эпимеризацией соответствующих кетоз и УДФ-альдоз. Реакции окисления алъдоз в
альдоновые кислотыты, дегидратации алъдоновых кислот до 2-кето-3-дезоксиальдоновых кислот и расщепления последних на соответствующие алъдозы и пируват
составляют пути распада углеводов у микроорганизмов.
Обмен карбоновых кислот и липидов. Обмен этих веществ тесно связан с обменом углеводов. Образующийся на предпоследней стадии гликолиза пируват в
результате окислительного декарбоксилирования превращается в ацетилкофермент А (АцКоА; см. Пантотеповая кислота). Т.к. АцКоА является непосредственным
предшественником в биосинтезе жирных кислот и изопреноидов, гликолиз и окислительное декарбоксилирование служат путем, в котором осуществляется превращение углеводов в
липиды. Наиболее распространенными липидами в организме животных являются триацилглицерины и некоторые изопреноиды. К последним относятся стероиды (главным
образом холестерин и его производные) и каротиноиды.
Обратный процесс - биосинтез углеводов из жиров - для животных не характерен. У растений и микроорганизмов он протекает в глиоксилатном цикле. В последнем
из образующегося в результате расщепления жирных кислот АцКоА синтезируется сукцинат, который в результате реакций окисления и декарбоксилирования
превращается в фосфоенолпируват. Далее из фосфоенолпирувата на амфиболическом участке пути гликолиза образуются углеводы.
Катаболический путь утилизации АцКоА состоит в окислении содержащегося в нем остатка уксусной кислоты в цикле трикарбоновых кислот до СО2 и
воды. При дефиците углеводов АцКоА для осуществления их биосинтеза образуется в результате расщепления жирных кислот или некоторых аминокислот. Таким
образом, у многих организмов цикл трикарбоновых кислот служит общим завершающим механизмом окисления углеводов, жиров и белков. В то же время у растений в
условиях фотосинтеза так называемый обращенный цикл трикарбоновых кислот может, подобно пентозофосфатному циклу, выполнять анаболическую функцию - превращать
CO2 в органические соединения.
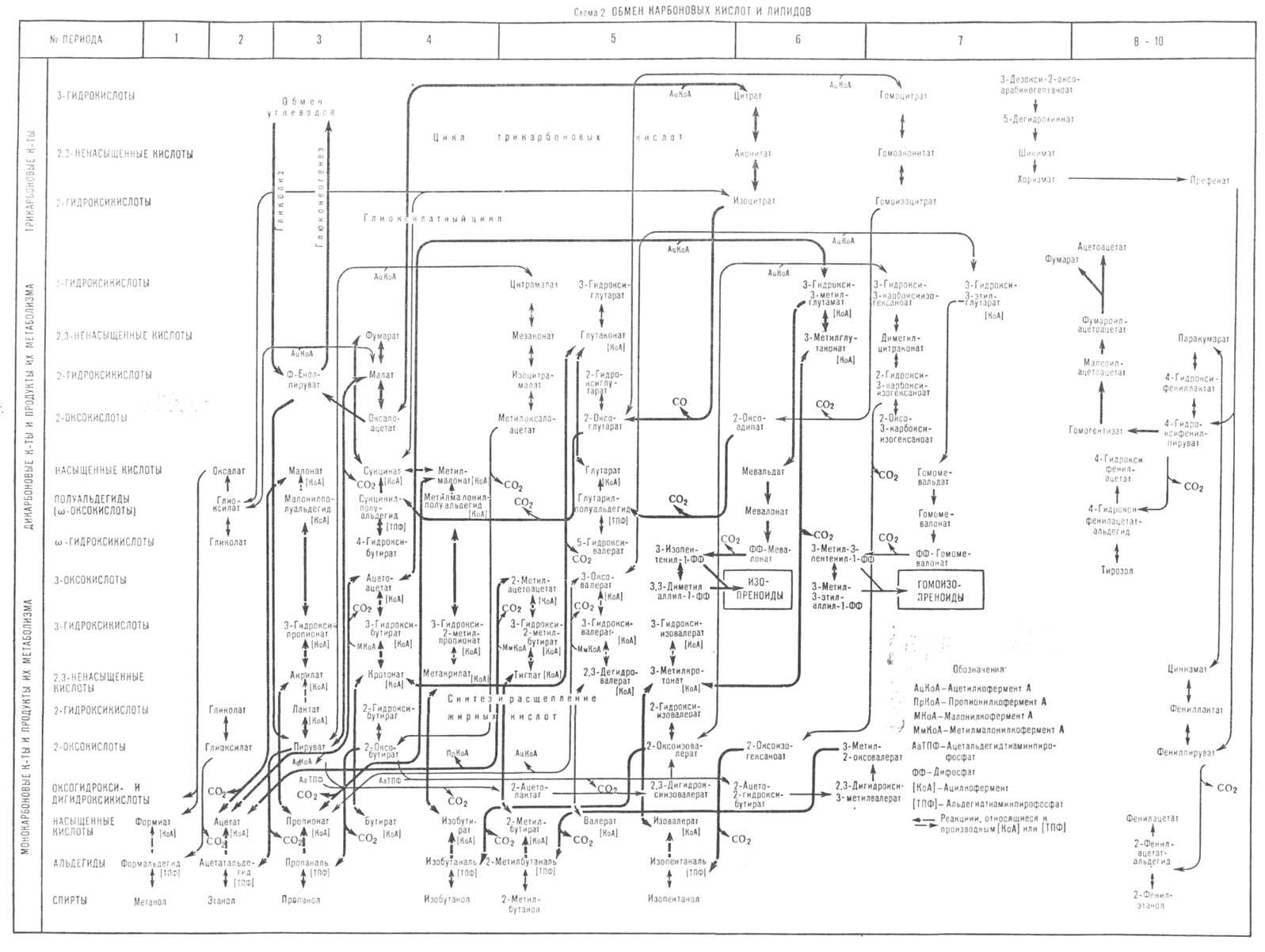
Основные метаболические пути в обмене карбоновых кислот и липидов приведены на схеме 2. На ней
показаны превращения три-, ди- и монокарбоновых кислот, а также их производных. Общим путем биосинтеза ди- и трикарбоновых кислот служат реакции конденсации
соответствующих моно- и дикарбоновых кислот с АцКоА или глиоксилатом. Три-, ди- и монокарбоновые кислоты переходят в соответствующие ди-, монокарбоновые
кислоты и альдегиды в реакциях декарбоксилирования. Связь между соединениями внутри периодов осуществляется в реакциях окисления и восстановления
карбонильных групп (соответственно до карбоксильных и гидроксильных), в обратимых реакциях дегидратации гидроксикислот в 2,3-ненасыщенные кислоты и
гидрирования 2,3-ненасыщенных кислот в насыщенные. В перечисленных реакциях образуются и расщепляются алифатические 2-оксокислоты - предшественники
соответствующих аминокислот. Углеродные скелеты оксокислот-предшественников валина и изолейцина (в периодах 5 и 6) - образуются в реакциях конденсации
соответственно пирувата и 2-оксо-бутирата с активным ацетальдегидом (продукт тиаминзависимого декарбоксилирования пирувата -
α-гидроксиэтил-тиаминопирофосфат; см. Тиамин). Реакции
оксопредшественников ароматических аминокислот представлены в периодах 7-10. Превращения дикарбоновых и трикарбоновых кислот в верхней части периодов 4-6
составляют цикл трикарбоновых кислот и глиоксилатный цикл. В нижней части схемы представлены начальные этапы путей биосинтеза и расщепления линейных и
разветвленных жирных кислот. Первые наращивают углеводородную цепь благодаря конденсации с ацетильным фрагментом малонил-кофермента А, переходя из периода
n в n + 2, вторые - в аналогичной реакции с пропионильным фрагментом метилмалонил-коферментом А, переходя из периода n в n + 3. В
периодах 6 и 7 приведены пути биосинтеза мевалоната и гомомевалоната - предшественников соответствующих изопреноидов (терпенов, стероидов, каротиноидов и
др.) и реже встречающихся гомоизопреноидов (например, ювенильный гормон насекомых).
Обмен азотсодержащих соединений. Первичный источник азота в обмене веществ - атмосфера. Непосредственно использовать свободный азот могут многие
виды бактерий. Однако большая часть микроорганизмов и все животные и растения усваивают лишь связанный азот в виде солей аммония, нитритов, нитратов или
продуктов расщепления белков. Основу внутреннего азотистого обмена составляют биосинтез и расщепление белков, нуклеиновых кислот и порфиринов. Аминокислоты в
организме образуются в реакциях восстановительного аминирования или переаминирования α-оксокислот. Белки включают лишь 20 из всех встречающихся
в живой природе аминокислот, называемых протеиногенными. Из них в организме высших животных синтезируется примерно половина. Другая половина относится к
незаменимым и в организм поступает только с пищей. Синтез полилептидной цепи белка из аминокислот (трансляция)осуществляется рибосомой. Последовательность аминокислот в белке определяется
последовательностью триплетов генетического кода в матричной РНК.
Катаболизм белков у всех организмов начинается с их расщепления по пептидным связям протеолитическими ферментами. В желудочно-кишечном тракте животных
белки гидролизуются трипсином, химотрипсином, пепсином и другими ферментами до свободных аминокислот, которые всасываются стенками кишечника и попадают в
кровоток. Часть аминокислот подвергается дезаминированию до оксокислот, претерпевающих дальнейшее расщепление, другая часть используется печенью или тканями
организма для биосинтеза белков. У млекопитающих отщепляющийся от аминокислот аммиак превращается в орнитиновом цикле в мочевину. Этот процесс осуществляется в печени. Образующаяся
мочевина вместе с другими растворимыми продуктами обмена веществ выводится из кровотока почками.
У человека половина всех тканевых белков расщепляется и строится заново в среднем в течение 80 сут, белки печени и сыворотки крови наполовину обновляются
каждые 10 сут, белки мышц-каждые 180 сут, отдельные ферменты печени-каждые 2-4 ч.
Нуклеиновые кислоты - продукты углеводного и азотистого обмена. ДНК образуется в клетке в результате репликации или обратной транскрипции из
дезоксирибонуклеозидтрифосфатов, РНК - в результате транскрипции из рибонуклеозидтрифосфатов. Нуклеиновые кислоты выполняют функции хранителей и переносчиков
наследственной информации, закодированной в последовательности четырех различных оснований в цепочках нуклеиновых кислот. Эта информация реализуется в
структуре белка. Через ферментативные и структурные функции белка она определяет наследуемые особенности обмена веществ организмов. Катаболизм
нуклеиновых кислот состоит в их гидролизе нуклеазами до нуклеотидов, которые затем расщепляются на составляющие их фосфат, пентозы, пуриновые и пиримидиновые
основания. При этом пуриновые основания окисляются до мочевой кислоты, которая у млекопитающих расщепляется до глиоксилата и мочевины. Пиримидиновые
основания в организме животных расщепляются до β-аланина и 3-аминоизобутирата.
Исходными субстратами в биосинтезе порфириновых соединений служат сукцинат и глицин. Порфириновые соединения выполняют в обмене веществ важные
функции, принимая участие в окислительно-восстановительных процессах. В частности, в составе гема в гемоглобине порфириновое кольцо участвует в переносе
О2 в крови. Порфириновое кольцо входит в состав цитохромов и хлорофиллов. Катаболизм порфиринов в животном организме состоит в раскрытии и
частичной деградации порфиринового кольца. Продукты катаболизма в виде окрашенных соединений (биливердина, билирубина и др.) наряду с продуктами частичного
окисления стероидов (холевыми кислотами) выводятся через желчные протоки в кишечник.
На схеме 3 показаны превращения пуриновых и пиримидиновых оснований, уреидов карбоновых кислот, а
также дикарбоновых и монокарбоновых аминокислот и их производных. Протеиногенные аминокислоты на схеме подчеркнуты (незаменимые для человека аминокислоты
отмечены звездочкой). Основной путь биосинтеза мочевины у млекопитающих (орнитиновый цикл) представлен в периоде 5. Реакции образования и расщепления
пуриновых и пиримидиновых оснований размещены в периодах 2-4 в верхней части схемы (для этих групп соединений периоды не всегда соответствуют количеству
атомов С в скелете молекулы). В средней части схемы показан путь биосинтеза предшественников порфириновых и корриновых соединений - d-аминолевулиновой
кислоты и порфо-билиногена из глицина и сукцината.
Взаимоотношение между обменом углеводов, липидов и карбоновых кислот, а также азотсодержащих соединений показано на схеме 4.
Схема 4. Обобщенная схема обмена веществ.
Минеральный обмен. Под минеральным обменом понимают процессы усвоения, превращения и выведения организмом неорганических веществ. Поскольку такие
неорганические вещества, как вода, СО2, NH3, фосфаты, сульфаты, иод и др., включаются при обмене веществ в органические
соединения, между органическим и минеральным обменами четкая граница отсутствует. Наибольший удельный вес в минеральном обмене занимает водно-солевой обмен,
в котором принимают участие катионы (Na+ , K+, Са2+ , Mg2+) и анионы (Сl-,
HPО42-, НСО3-, SO42-). В результате активного трансмембранного переноса ионы Na+
непрерывно удаляются из клеток в межклеточную среду, а замещающие их ионы К+ концентрируются внутри клеток. Ионы Са2+ у животных
участвуют в проведении нервного импульса, поэтому постоянство их концентрации в организме имеет существенное значение для нормального функционирования
нервной системы. У позвоночных животных Са2+ и фосфат НРО42- в виде кальциевых солей фосфорной кислоты служат главными
компонентами костной ткани. Из карбонатов Са и Mg беспозвоночные строят раковины. У растений Mg2+ входит в состав хлорофилла. Сульфатная группа -
фрагмент желчных кислот и хондроитинсульфата хрящей. В организме сульфат образуется в результате окисления сульфгидрильных групп цистеина и гомоцистеина.
Водно-солевой обмен обеспечивает постоянство ионного состава, осмотического давления, рН и объема жидкостей организма. Потребность взрослого человека в
Na+, K+, Са2+, Mg2+, Сl- и НРО42- составляет соответственно 215, 75, 60, 35, 25
и 105 мг/сут. Выведение из организма растворимых минеральных продуктов обмена веществ, так же как и органических, осуществляется у человека и животных
почками.
Функция многих металлов (например, железа, меди, цинка, кобальта, молибдена) в обмене веществ состоит в их участии в биохимических реакциях в
качестве коферментов. Иод участвует в обмене веществ в составе гормона тироксина. Большая часть из перечисленных элементов присутствует в организме в
концентрации меньшей, чем 10-3%, и относится к микроэлементам. Особо в обмене веществ рассматривают минеральное питание растений. Соли
аммония, калия, нитраты, фосфаты и микроэлементы широко используют в качестве удобрений, улучшающих рост и увеличивающих продуктивность растений.
Энергетический обмен
Движущей силой обмена веществ служит разность термодинамических потенциалов участвующих в реакциях соединений. По источникам используемой при
обмене веществ энергии организмы делят на фототрофы и хемотрофы. У первых роль богатых энергией субстратов выполняют эндогенные продукты начального
этапа фотосинтеза, у вторых - различные соединения экзогенной природы. Если в качестве субстратов окисления хемотрофы используют органические соединения, то
их относят к хемоорганотрофам, если неорганические (например, Н2, NH3, CO, Н2S) - к литотрофам.
В рамках общего обмена веществ изучение энергетического обмена у фототрофов состоит в определении энергетических параметров фотосинтеза, у
хемотрофов - в исследовании баланса между энергией, выделяемой при расщеплении субстратов обмена веществ, и энергией, расходуемой на биосинтез
конечных продуктов обмена веществ, на совершение механической работы, а также рассеиваемой в виде тепла. В общем энергетическом обмене животных
выделяют основной обмен - минимальное количество энергии, необходимое для поддержания жизни организма в состоянии покоя. У взрослого человека он составляет
1600-1700 ккал/сут (6700-7100 кДж/сут). Количество тепла, выделяемое теплокровными животными при основном обмене, пропорционально поверхности их тела
(правило Рубнера). Более точно теплопродукция (в ккал/сут) организма животных выражается формулой W = 70 М0,75 (М-масса тела в кг). Данные об
общем энергетическом обмене используют для вычисления калорийности продуктов питания и составления сбалансированных по энергозатратам организма рационов
питания.
Молекулярные механизмы генерирования и утилизации энергии на промежуточных этапах обмена веществ изучает биоэнергетика, которая рассматривает сопряжение биологического окисления с
фосфорилированием. Это обусловлено тем, что свободная энергия гидролиза основного продукта фосфорилирования - АТФ и в меньшей степени других фосфатных производных, например гуанозинтрифосфата,
креатинфосфата, - обеспечивает в сопряженных реакциях синтез сложных соединений, мышечное сокращение, транспорт соединений через биологические мембраны
против градиента концентрации (активный транспорт), создание на мембране электрического потенциала, разряд которого, в частности, обеспечивает проведение
нервного импульса и другие биоэлектрические явления. Энергия гидролиза АТФ может также трансформироваться в световую энергию или служить в организме
источником тепла.
Различают субстратное фосфорилирование, состоящее в фосфорилировании молекул субстрата при их окислении никотинамидными коферментами (см. Ниацин) и окислительное фосфорилирование, наблюдаемое на стадии образования АТФ из
аденозиндифосфата (АДФ) при переносе электронов (атомов водорода) по цепи коферментов в процессе дыхания и в фотосинтезе. Пример субстратного
фосфорилирования - катализируемая в гликолизе глицсральдегидфосфат-дегидрогеназой реакция образования D-1,3-дифосфоглицери-новой кислоты
(НО)2Р(О)ОСН2СН(ОН)С(О)ОР(О)(ОН)2 из неорганического фосфата и D-глицеральдегид-3-фосфата. Фосфатная группа в положении 1
этого соединения с помощью фермента фосфоглицераткиназы может переходить к АДФ, образуя АТФ. Субстратное фосфорилирование более характерно для организмов,
живущих в анаэробных условиях. Объектом для переноса электронов (атомов водорода) у них обычно служат карбонильные группы промежуточных продуктов окисления
субстрата. Так, НАДН (восстановл. форма никотинамидадениндинуклеотида) восстанавливает продукт декарбоксилирования пирувата (ацетальдегид) до этанола (при
брожении), а пируват - до лактата (при гликолизе). Пример окислительного фосфорилирования - синтез АТФ в аэробных условиях, а также у сульфатных бактерий в
анаэробных условиях. Во всех этих случаях образование АТФ сопряжено с переносом электронов (атомов водорода) по цепи окислительно-восстановительных
коферментов, включающей никотинамидные и флавиновые коферменты (см. Рибофлавин), убихиноны (у животных), пластохиноны (у растений), негеминовое
железо и медь. В митохондриях первонач. донором электронов (атомов водорода) служат органические соединения, а конечным акцептором - атмосферный
О2, в хлоропластах донором и акцептором электронов (атомов водорода) служат продукты фотореакции. У сульфатных бактерий донором являются
органические соединения, а акцептором-сульфат, восстанавливаемый до сульфида.
Особенность биологического окисления в аэробных условиях состоит в том, что органический субстрат обычно полностью окисляется до СО2 и воды в
циклической последовательностях реакций. Пример такого окисления - превращения в цикле трикарбоновых кислот ацетата, образующегося в виде АцКоА при
окислительном расщеплении жирных кислот, углеводов и некоторых аминокислот. Другой пример - полное окисление глюкозы в пентозофосфатном цикле. Циклические
пути благодаря полному окислению субстратов позволяют извлекать из органических соединений максимум заключенной в них свободной энергии.
Регуляция обмена веществ
На обмен веществ постоянно оказывают воздействие различные факторы внешней и внутренней среды. Большая часть из них эффективно используется
организмами для своего роста и развития. Это происходит благодаря функционированию механизмов регуляции обмена веществ. Наиболее простым из них,
способствующим сохранению внутренней среды организма (поддерживанию гомеостаза), является механизм восстановления в химической системе равновесия в
соответствии с законом действующих масс. Благодаря этому значения рН в буферных жидкостях организма устойчивы к случайным воздействиям. Предотвращение
накопления в организме невыводимых продуктов обмена веществ также осуществляется благодаря восстановлению равновесия в замыкающих участках циклических
путей обмена веществ.
Более сложные механизмы регуляции обмена веществ обусловлены прямыми и обратными управляющими связями. Суть их состоит в воздействии метаболитов на
интенсивность биохимических процессов, в которых они сами образуются или испытывают превращения. В обмене веществ регуляция активности ферментов часто
осуществляется посредством аллостерического взаимодействия ферментов с субстратами или промежуточными продуктами (см. Ферменты). Классический пример подобной регуляции с отрицательной обратной связью -
подавление изолейцином собственного биосинтеза в результате его аллостерического взаимодействия с ферментом треониндегидратаза, катализирующим начальную
реакцию пути биосинтеза изолейцина. Пример положительной прямой связи - стимуляция синтеза фосфоенолпирувата в гликолизе предшествующими метаболитами:
фруктозо-1,6-дифосфатом, глюкозо-6-фосфатом и глицеральдегид-3-фос-фатом. Управляющие связи такого рода позволяют стабилизировать концентрации метаболитов в
неравновесной системе.
Сходным образом осуществляется регуляция обмена веществ на уровне биосинтеза ферментов. При этом субстрат или продукт реакции регулирует активность
белкового репрессора, подавляющего транскрипцию (синтез матричной РНК на ДНК-матрице) соответствующего оперона (участок ДНК, кодирующий одну молекулу
матричной РНК под контролем белка-рспрессора). Примером регуляции при помощи положительной прямой связи может служить в данном случае управление расщеплением
лактозы. Появление в среде лактозы инактивирует у бактерии Escherichia coli соответствующий репрессор и тем самым разрешает транскрипцию оперона,
кодирующего ферменты, катализирующие расщепление лактозы. Пример регуляции при помощи отрицательной обратной связи - управление биосинтезом гистидина.
Избыток гистидина активирует репрессор, ингибирующий транскрипцию оперона, кодирующего ферменты биосинтеза гистидина. Если репрессор и белки, синтез которых
он подавляет, кодируются одним опероном, то отрицательная обратная связь осуществляется без участия внешних модуляторов активности репрессора. Аналогичным
образом осуществляется регуляция биосинтеза белка на уровне трансляции (синтез белка на РНК-матрице). Такой механизм регуляции позволяет синтезировать белок
в строгом соответствии с потребностью в нем на данном этапе существования организма.
Многоклеточные организмы наряду с рассмотренными внутриклеточными механизмами имеют надклеточные - гормональные механизмы регуляции обмена веществ.
Гормональная регуляция координирует обмен веществ в различных тканях и органах и интегрирует его в рамках организма в целостную систему. Гормональная
регуляция обмена веществ у растений осуществляется группой фитогормонов, например ауксинами и гиббереллинами. Гормональную регуляцию обмена
веществ у животных осуществляет эндокринная система, источниками гормонов в которой являются центральные и переферические железы внутренней секреции.
Характер управляющих связей в этой системе иллюстрирует механизм поддержания концентрации глюкозы в крови на постоянном уровне. Так, повышение концентрации
глюкозы в крови увеличивает продукцию инсулина, который стимулирует клетки на усиленное потребление глюкозы. Возникающий при этом дефицит глюкозы приводит к
увеличению продукции другого пептидного гормона - глюкагона, который стимулирует восстановление концентрации глюкозы благодаря расщеплению гликогена в
клетках.
Как правило, механизмы гормональной регуляции многоступенчаты. Воздействие гормонов на обмен веществ осуществляется через клеточную мембрану, во
многих случаях посредством активирования аденилатциклазной системы (см. Аденилат-циклаза). Обратные связи в эндокринной системе часто замыкаются через
нервную систему. При этом нервная система, получая сигналы из внешней среды или от внутренних органов, управляет железами внутренней секреции. например,
гипоталамус по сигналам от центральной нервной системы, передаваемым гормонами-медиаторами (например, норадреналином, ацетилхолином), секретирует пептидные
нейрогормоны (релизинг-факторы), разрешающие секрецию гормонов гипофиза. Последние стимулируют секрецию гормонов периферическими эндокринными железами. Эти
гормоны влияют на обмен веществ в соответствующих органах и тканях таким образом, чтобы компенсировать изменения во внутренней среде или подготовиться
к возможным ее изменениям, прогнозируемым центральной нервной системой (например, при стрессовых ситуациях). Гипоталамо-гипофизарная система, в частности,
играет центральную роль в регуляции водно-солевого обмена животных (см. Вазопрессин, Окситоцин).
Регуляция при помощи управляющих связей допускает возникновение в обмене веществ состояний с автоколебательными режимами (см. Колебательные реакции), обусловливающими периодическое изменение концентраций
некоторых метаболитов. Такие автоколебательные режимы лежат в основе различных периодических процессов у живых организмов, например сердечных сокращений,
суточных ритмов активности и др.
Нарушения обмена веществ
Различают нарушения обмена веществ, вызываемые прямым влиянием на него неблагоприятных факторов (недостаток или несбалансированность органических и
минеральных субстратов обмена веществ, избыточности или недостатка внешних физических воздействий - температуры, света, звука и др.), и нарушения,
вызываемые изменениями в наследственном аппарате организма. В соответствии с этим заболевания обмена веществ у людей подразделяют на болезни
недостаточности и врожденные болезни. например, к болезням недостаточности относятся алиментарная дистрофия при голодании или недостатке в пище незаменимых
аминокислот, авитаминозы (цинга - при недостатке аскорбиновой кислоты, рахит - при недостатке витамина D). К болезням недостаточности более чувствительны
организмы в стадии роста и развития. Примеры врожденных болезней - алкаптонурия, связанная с недостаточной активностью ферментов, расщепляющих
гомогентезиновую кислоту, и серповидноклеточная анемия (см. Гемоглобин).
Нарушения обмена веществ у микроорганизмов, вызванные изменениями в составе субстратов или полученные в результате мутагенеза, широко используют в
практических целях. Так, добавляя в питательную среду дрожжей сульфит натрия, удается переключить алкогольное брожение на глицериновое и создать на этой
основе биотехнологию получения глицерина. В микробиологической промышленности широко используют полученные селекцией штаммы микроорганизмов - суперпродуценты
отдельных аминокислот, антибиотиков и др. Методы генной инженерии позволяют избирательно изменять наследственный аппарат клеток и благодаря этому
целенаправленно воздействовать на структуру и динамику обмена веществ у организмов.
Лит.: Дэгли С., Никольсон Д., Метаболические пути, пер. с англ., М., 1973; Ньюсхолм Э., Старт К., Регуляция метаболизма, М., 1977; Лукнер М.,
Вторичный метаболизм у микроорганизмов, растений и животных, пер. с англ., М., 1979; Мак-Мюррей У., Обмен веществ у человека, пер. с англ., М., 1980; Малыгин
А. Г., Симметрия сети реакций метаболизма, М., 1984.
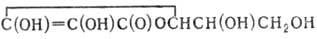
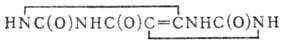
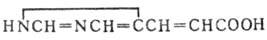
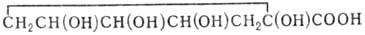
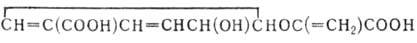
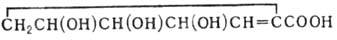
{kind=link}
{kind=link}
{kind=link}